|
|
|
||||||||||||
|
|
|
||||||||||||
|
|
|
|
|||||||||||
|
|
|
|
|||||||||||
|
|
|
|
|||||||||||
|
|
|
||||||||||||
|
|
|
|
|
||||||||||
|
|
The Transition from Hominids to Hominins
Clickable terms are red on the yellow or green background |
|
|||||||||||
|
|
The
Theoretical Foundations of Anthropology, Ethnology and Prehistoric
Studies |
|
|||||||||||
|
|
|
|
|
||||||||||

The
Transition from Hominids to Hominins
Similar lineages have to be traced up also
for hominins and recent racial varieties of humankind. The principal dichotomy
splits them into the Negritic herbivorous (pre)agriculturalists in the
tropical equatorial zone and the Altaic carnivorous (pre)pastoralists in
colder regions of Current population genetics defends African origins and Chris Stringer’s ‘single-origin model’. It supports theories
of strict monogenism in belief that one sapient race colonised the world from
one homeland in
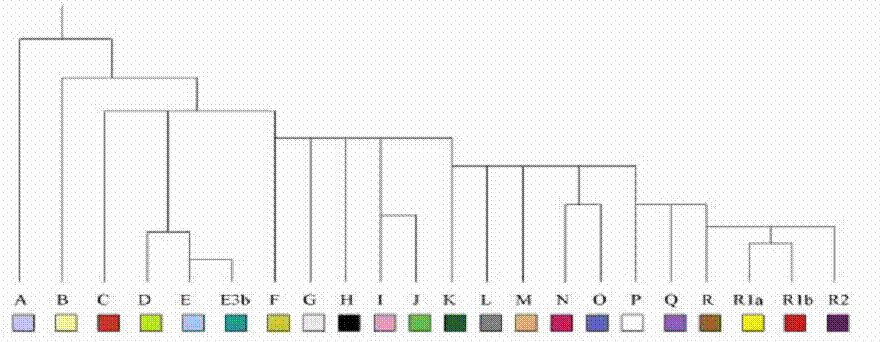
Table 5.
The simplified genealogic tree
of Y-DNA haplogroups The out-out-Africa wandering sounds like
as tenable hypothesis but its dating must be shifted to the earlier times of
Oldowan migrations from 1.8 mya to 0.5 mya. There exists little evidence
supporting the alternative theory of an out-of-Asia model but it is worth taking into account. Besides the
Riwat flake-tool culture (2 mya) in north Pakistan it is necessary to
scrutinise Asiatic hominins as an independent genetic strain evolving
inclinations to green, yellow and red pigmentation. Asiatic orangutans and
gibbons exhibited green, grey, yellow and red hair pigmentation. Prehistoric
megalith-builders in
Table 6.
The Y-DNA transitions during the out-of-Africa and
out-of-Asia exodus Palaeontological digs are relatively rare,
so their taxonomic labels have to be determined in accordance with other
prehistoric disciplines. The categories of their diachronic phylogenesis may
be reconstructed by comparison to synchronic raceology and the ethnology of
living varieties. Another guideline is found in the typology and distribution
of archaeological cultures because their bearers were ancient anthropological
and ethnic groups. Table 6 proposes to solve their mutual correspondences and
structural resemblances like an equation in several unknowns. The evolution
of human races gets clear contours if their isolated types are arranged into
migratory chains linked by similar cultural patterns. The Balkan Dinaric race
can be related to the mythological breed of the Mycenaean Cyclopes and the
builders of Megalithic monuments in |
|
Table 7. Joint lineages of Palaeolithic and Neolithic
races Comparative parallels confirm that humans
races are not isolated local outgrowths but lawful descendant leftovers of
one prehistoric migration. Cultural comparison witnesses that African Negrids must have developed into Oceanic Melanesids,
Australids and American Amazonids. Despite certain minor differences the
South African Sanids seem to be related to Pygmies, European Lappids and
Asiatic Sinids. Leptolithic technology betrays hidden affiliation between
stocks of the Kafrids, European Pelasgids and Asiatic Tungids. Surprising
coincidences associate the African Khoids and Mousterian Neanderthalers, who
both excelled in big-game hunting, beehive architecture and manufacturing
leaf-shaped heads of lances. The most astonishing phenomenon about such
chains of prehistoric relatives appears in similarities of their language
structures.
Obvious genetic links between remote continental stocks call for an
integrated anthropological reclassification. The first hominids born in Palaeo-races, proto-races and
deutero-races formed archetypal stocks of humanity, who influenced each other
and intermarried as free genetic strains. They represented pure clear-cut
somatic taxa with definite ethnonymy, archaeological industry, religious
beliefs and cultural customs. The modern living racial groups are hardly more
than their mixed and degenerate residual derivates and have to be kept apart
as subsequent ulterior neo-races. An analogous subcategorisation has
to be carried out also in ethnology and comparative linguistics. These fields
deal only with neo-tribes and neo-languages and bury their
earlier predecessors as dead and extinct. Their real goal is to decompose
them into original pure typological elements arisen from palaeo-tribes,
palaeo-cultures and proto-languages. The Palaeolithic Survival Theory1 assumes that it is possible to
reconstruct their origins in as lawful integral genealogies as are presently
ruling in palaeoanthropology and archaeology. Such phylogenetic trees of racial varieties partly agree with
genealogies looming in population genetics. Sanids exhibited the Y-DNA
haplogroup A, the Khoids spread the Y-DNA haplogroup B, the Negrids the
haplotype E, and the Kafrids propagated the Y-DNA haplogroups T or C. DNA
haplogroups serve as an efficient tool of investigating racial interrelations
but their scope of study is confined to the last 70, 000 years. This is why
they fail to give a convincing account of earlier prehistory. Historical
linguistics is accustomed to work with ‘short-range comparisons’, while they
provide efficient methods for enquiring into ‘middle-range comparisons’
between prehistoric stocks. However, it is only archaeology and
palaeoanthropology that make it possible to plot ‘long-range genealogies’.
Generally speaking, neo-races, neo-tribes and neo-languages are only
fragmentary splinters fallen away from the mainstreams of Palaeolithic
anthropogenesis. Besides distilling into pure subcomponents they need
rearranging into regional subclades and lineages. Every Palaeolithic
palaeo-race and palaeo-tribe is an octopus spreading its tentacles radially
in several dominant directions and so their convenient subclassification
partitions racial varieties according to regional lineages. The
dominant super-lineage unites the regional outgrows of the black equatorial
races. It is fathered by its progenitor Homo erectus and its migratory
plantations should be kept together by a common taxon of Negrids or
Congids. Its continental tentacles ought to be denominated as Euro-Negrids,
Levanto-Negrids, Georgo-Negrids, Indo-Negrids, Sino-Negrids, Australo-Negrids
and Australo-Negrids. Their unity with the ancestral stock of Bantu people in
Extract from P. Bělíček::
The Synthetic Classification of Human
Phenotypes and Varieties Prague
2018, pp. 19-21 |
||||||||||||||||||||||||||||||||||||||||||||||
|
The Paragenetic Model of Human Evolution from
Hominids The paragenetic model of prehistory presupposes
that most hominins and hominids lived in relative interbreeding and their genetic
distances were much nearer than now. What we denote as detached genera and
species were actually interfertile genetic races, strains, lineages, crosses
and hybrids that later lost mutual interfertility owing to isolation in
different hominoid populations. It is not plausible that they developed by
large jumps from one genus to another, they must have maintained and
preserved their genetic pool through progressive evolutionary metamorphoses. Many
categories of genus and species were only generations, so the extant binomial
and trinomial anthropological classification should adopt a special term for
transient generations. All strains underwent parallel processes of
hominisation, gracilisation and sapientisation by means of radical
revolutions and longer stages of conservative inertia. Hominins split off
hominids and hominoids as a special genetic stream competing with alternative
strains of Paranthropines and Australopithecines. Participation in different
population strains caused intraspecial differentiation. In the following
evolutionary series the symbol means digression while the arrow → implies genetic continuity. It does not
mean direct mother-daugher inheritance but a complex statistic process with
many digressions splitting off the dominant mainstream. The following series
are chief statistic mainstreams that suggest that the Palaeolithic Urrassen
had different ancestors but converged to one of predominant Neolithic racial
varieties. Tall robust
dolichocephalous herbivores with marked crista sagittalis Gigantopithecus
(9 mya) → Ouranopithecus (9 mya) (
Gorillas (9 mya)) → Paranthropus aethiopicus
(2.5 mya) ( Paranthropus robustus (2
mya)) → Australopithecus garhi
(2.5 mya) → Australopithecus sediba (1.8 mya) → Homo gautengensis (1.8
mya) → Homo erectus
(1.8 mya) → Oldowans (1.8 mya). Slender
piscivores with tall and leptoprosopic flattish faces: Proconsul africanus (23 mya) → Kenyanthropus platyops (3.5 mya) → Australopithecus afarensis (3.9–2.9 mya) → Homo
habilis (2.1–1.5 mya) → Homo rudolfensis (2–1.5 mya) →
Levalloisians (0.5 mya). Tall
brachycephalous carnivores and big-game hunters with narrow aquiline noses Australopithecus anamensis (4.5 mya) → Laetoli man → Homo heidebergensis →
Homo rhodensis (0.5 mya) ( Saldanha
man) →
Homo neanderthalensis → Mousterians. Shortsized brachycephalous omnivores: Ardipithecus ramidus (4.4
mya) → Ardipithecus kadabba ( Pan
paniscus (Bonobo)) → Australo-pithecus afarensis (3.9 mya) → Homo habilis →
Sanids → Pygmids ( Homo floresiensis) → Sinids.
Table 1. The paragenetic model of racial
diversification |
|
|||||||||||||||||||||||||||||||||||||||||||||||