|
|
Negrids: Plant-Gatherers with Pebblestone Choppers and Rectangular Huts
Traditional accounts of the black people’s
anthropogenesis reckon with late dating and derive their origin from the
Early Neolithic Asselar man. He lived around 6,400 BP and his fossil remains were found in the massif Adrar des Ifoghas in Mali. This Early Neolithic
predecessor purportedly spoke the common Bantu language and acted as the
ancestor of all Bantu people. Classic Africanist
philological studies counted with even more recent dating. Their founder Harry H. Johnston thought that the Common Bantu unity came into
being soon after the beginning of our era. Such estimates, however,
contradict the available archaeological evidence. A great number of
archaeologists believe that the Bantu origins may be traced back to the Sangoan culture (130,000 BP). It reigned in the territory of Central Africa and produced innovated types
of hand-axes, picks and pebble-stone choppers. Its successor later appeared
in the Lupembian hand-axe tradition (40,000 BC)
centred in the Congo rainforests. Most
archaeologists associate them with Acheuleans roots
and cultural patterns drifting from Arabic countries. Other authors classify Sangoan advances as an autochthonous development of Oldowan plant-gatherers. This view is supported by the
distribution of Sangoan finds in the Congo considered as the homeland
of Bantu people and African Negrids.
The mainstream of prehistoric studies
supports theories of unilinear hominisation,
sapientisation and gracilisation
that see the very champion of humanity in Homo sapiens. They suppose
that this human ancestor developed from Homo erectus in East Africa and originated as early as 315,000 years
ago. Counterarguments object by emphasising the typological viewpoint and the
criteria of structural consistence. The Oldowan, Acheulean, Sangoan and Lupembian cultures continued consistently in the wake of Homo
erectus and his axe-tool manufacturing industry. These cultures preferred
vegetal sustenance and marched in the track of Cliff J. Jolly’s
vegetarian robust seed-eaters represented by Paranthropus
robustus and Paranthropus
boisei. On the other hand, the East African Homo
sapiens represented an alternative line of the Levalloisian flake-tool
tradition dated to 125,000 or even 500,000 years ago. His distribution was
confined to lakeside districts in the East African Depression connected to
northern sunken areas as far as the Jordan Rift in the Levant. He probably evolved from Raymond A.
Dart’s osteodontoceratic culture. This line was initiated by the gracile carnivore Australopithecus africanus
that
produced tools out of bones,
teeth and horns. His hordes lived on nomadic fishing and sought
shelters in caves. The empty
period between their eras was probably filled up by the bloom of Homo habilis, who was an antipode of plant-gathering
herbivores with vegetal subsistence. Anthropologists were fascinated by his
gentle and gracile countenance and glorified him as
a predecessor of Homo sapiens. Such biased prejudices overrated facial morphology and
condemned Homo erectus
to extinction. They forgot that gracile features
were often due only to lesser sexual dimorphism and there were several roads
inciting rapid mental development. Besides exerting manual skills it is
necessary to take into consideration short-term mutations and adapting to the
local climate.
Much confusion is caused by insisting on
strict monogenism and disregarding independent
parallel streams of sapientisation in Europe and Southeast Asia. Erroneous theories of linear gracilisation,
one-way sapientisation and single-origin
hominisation deepen the abyss
gaping between the allegedly extinct archaic genera and modern living races.
They hinder from grasping the genetic unity of all Negrids
in Jolly’s ‘robust herbivores with axe-tools’, who
spread all over the world by Oldowan colonisations.
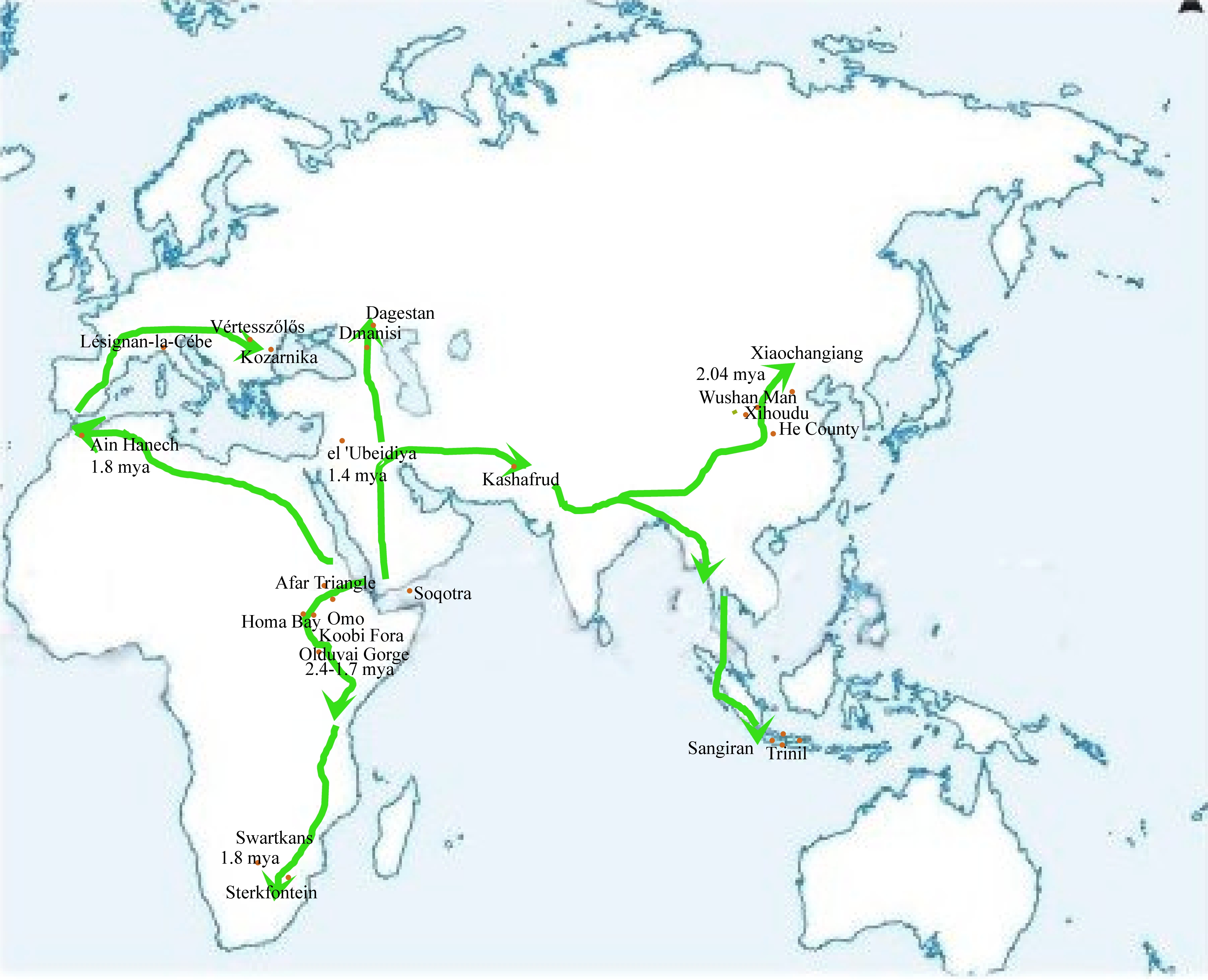
This diaspora was triggered by the eastward
propagation of Homo erectus and his expansion from
Africa to the Near East. Depots of Oldowan pebble-stone choppers were discovered in India, China, and Java. Excavations of
the Hoabinhian and Bacsonian
culture in Vietnam proved that the specific
technology of their knapping flourished as late as the Pre-Neolithic horizon.
Archaeologists assume that Homo erectus began to spread Oldowan pebble-stone chopper industry cca
1.9 million years ago and his cultural
mission ended as late as 10,000 years before present. His propagation naturally did not immediately span
as far as all remote localities of the present-day distribution,
the first African exodus created a
secondary homeland on Southeast Asia and its
progeny later continued travels to Australia and Melanesia. Map 2
demonstrates his prehistoric travels to Africa to the Levant, the Caucasus, Indochina and Melanesia. Hypothetical
reconstructions reckon also with migrations to Europe, Australia and the New World.
Homo
erectus was
buried alive although his archaeological sites with pebblestone
chopping tools clearly survived until 3000 BC. Deposits with unifacial choppers were discovered in the Hoabinhian and Bacsonian
cultures extending from Vietnam to Thailand, Malaysia, Java and Borneo. Wherever his hosts marched,
there emerged plantations of the black equatorial race preserving the
cultural customs of remote African ancestors. Their most characteristic
common feature consisted in vegetal subsistence with preagricultural
plant-gathering dispositions that have later led to the invention of field
cultivation. Traditional integralist approaches
deny his clear-cut typological character: they mix plant-gatherers with
hunters and fishers and his axe-tools for unearthing vegetal roots with
flint-flake projectiles. His most conspicuous distinguishing marker surviving
to our days is that both sexes wear fringed grass aprons and adult women go
out barebreasted with naked top. In his societies
women play a dominant constitutive role and hold a privileged position in
social affairs. His society preserves marital endogamy, matrilineal
inheritance and matrilocal marriage. Husbands live
in their wives’ place in large collective families and inhabit their wooden
rectangular longhouse with two-slope roofs.
|
|
Ethnic and cultural customs prove to be as perennial as somatic and
anatomic features. Table 23 sums up the common traits of all Negrids into paradigmatic patterns based on comparative
methods of typological generalisation. They include also typical markers of
anthropological phenotypes such as tall robust stature, dolichocephalous
skulls, high nasal indices and chamaerrhinia marked
by broad noses. Further common symptoms concern dark black skin and woolly
curly hair. Their systematic survey is completed by data of population
genetics attesting the existence of one homogeneous race.
|
Ecotype
|
lowlands with fertile
alluvial soil suitable for rich plant vegetation
|
|
Nutrition
|
vegetarian herbivorous and granivorous dispositions, plant-gathering economy,
shifting and fallow agriculture, slash and burn farming
|
|
Dwelling
|
rectangular longhouses for
large families with two opposite half- timber buildings facing one another
and living in matrilocal endogamy
|
|
Cult
|
ancestral cults, endophagy, naturistic
hylozoistic polytheism revering primordial elements of air, earth, water
and fire, priests employed as rain-makers, matriarchal organisation,
chthonic myths
|
|
Burials
|
interment under the kitchen
floor or on head-benches for sleeping
|
|
Death
|
ancestral cults adore
forefathers as gods, keep them in nearest proximity and try to identify
with their remains by ritual endophagy, i.e.
burial feasts with eating their dead corpse and drinking their blood
|
|
Visage
|
tall robust and muscular
stature with a dolichocephalous cranium, chamaerrhinia
with broad noses and high nasal indices, dark skin, dark woolly and curly
hair, brown eye colour
|
|
Industry
|
Oldowan pebble-stone chopping-tools, heavy choppers
|
|
Clothing
|
short fringed grass
aprons, women go out with naked breasts
|
|
DNA
|
ABO group O, Rh+, Y DNA E, K, M, mtDNA L →
mtDNA H
|
|
Languages
|
initial prenasalised stops mb-,
nd-, ng-, voiced-to-surd
consonant correlation, prefixing nominal morphology, nominal classes and
classifiers, animate b-plurals, SVO word-order, subordinate that-clauses
instead of participles and gerunds
|
Table 23. The cultural paradigm of Oldowan plantations
Such typological parallels contradict the traditional recent dating
that does not correspond to the real chronology of Homo erectus and
his Oldowan finds. They uproot the inveterate dogmas claiming that there was
only one gracile omnipotent and omniscient Homo sapiens that
exterminated and aryanised other inferior species. Evolution promotes
ascendant progress in all continental races and enhances their vitality by
mutual miscegenation. It conditions ascendant growth by perennial genetic
inheritance and rejects fallacious Eurocentrism. It also impugns the idea
that one homogeneous Nostratic and Indo-European nation could split into
different races in a few centuries. Palaeolithic races were prior to
Neolithic ethnic tribes and the latter were prior to nations of the Late
Middle Ages.
Extract from Pavel Bělíček The Synthetic Classification of Human
Phenotypes and Varieties, Prague 2018, pp. 69-75
|
|
|
|
The Subdivision of African Negrids
Archaeological evidence splits African Negrids
into three principal generations: (1) Oldowans (2.5
mya), (2) Sangoans
(130,000 BP) and (3) Lumpembans (40,000 BP). Oldowans prevailed in East Africa, the latter two
concentrated in the tropical rainforest area. These cultures seem to act as
main milestones in the transition from plant-gathering economy to field
cultivation and the development of African Neolithic agriculture. The
Neolithic hand-axes served to the early farmers as mattocks and hoes for
uprooting vegetal roots and their achievements achievement exhibited very
small progress for detailed periodisation. It is
probable that Oldowans have survived as Palaeo-Negrids while modern Negrids
descend from Sangoans.
Palaeo-Negrids.
In isolated refuges of Africa, Europe and Asia it is possible
to come across archaic populations reminiscent of the earliest settlers that
found their homes in Europe at the dawn of the Lower
Palaeolithic Age. The French anthropologist George Montandon subsumed their group
as Homo s. palaeniger. They were notable for bluish black
skin colour, deep-set eye sockets and quadrangular eye-holes. Further characteristic
features were heavy brow arches, heavy jaws, sloping foreheads and receding
chins.
European Palaeo-Negrids. The first European colonists were a
progeny of Homo erectus (Tautavel man in France) and Homo antecessor (Atapuerca man)
and came to Europe with a host of wanderers
via Gibraltar. Their people were later superseded by the Acheulean hand-axe makers, who arose by mixing the
African Negroid heritage with prefixing classifiers and the Levallosian people, who spoke a language of agglutinating
type. They probably came from Aden in Saudi Arabia and via Levant and Anatolia they arrived in
the Balkans and Central Europe. In western Europe they met
remains of Homo antecessor, who profiled as
a Negrid with less sophisticated hand-axe industry
of Chellean or Abbevilian
style. His progeny was assimilated by Acheulean
newcomers but may have survived partly in the Old Black Breed. Their variety
was discovered by W. Ripley in the Shetland Islands between
1897 and 1898. In Scandinavia it
appeared as the Tydal race, whose dialects apply prenasalised stops.
Zambesids. The Scottish Old Black Breed exhibited deep-set eyes,
heavy brow ridges and further archaic tendencies similar to African Palaeo-Negrids. Their proto-type was classified by George
Montandon as Homo s. palaeniger. His characteristic features were deep-set eyes,
rectangular eye-holes, heavy brow arches, heavy jaws, very wide faces,
sloping foreheads, receding chins, strong hairiness and bluish black skin
colour. Such tendencies appeared inconsistently also in South and Central
Bantuids but they were
prominent especially among the tribes of Kwanyama,
Hlubi, Fengu and Makua. In condensed form they cropped up in the racial
varieties of Katangids, Bergdama
and Shara tribes. H. V. Vallois
proposed to class them as a special type of Zambesids. H. Vedder
gave preference to the term of Bergdama for their peculiarities. B. Lundman was fascinated by the bluish back skin colour in
the Shara tribes and suggested to call them Sharids.
The current results of
population genetics imply that the earliest ancestors concentrated around the
original homeland laying somewhere in Cameroon
and lower reaches of the Congo.
Its area was originally populated by the Palaeo-Negrids
with the Y-haplogroup E*-M96 and the mt-haplotype L0. Yet their settlements were later
overlaid by Neo-Negrid incomers producing the Sangoan industry (130,000 BP) with a more sophisticated
axe-tool industry. They superseded Palaeo-Negrids
in their old heartland and now coincide with the racial group of Congids. The next step brought differentiation
between Bantuids prevailing in East Africa from Somalia
to Swaziland. The term of Bantu is not the original ethnonym of all African blacks, its word root is prefixed
by the plural classifier ba- attached to
plural multitudes of humans. Its
root -ntu must indicate the tribes of Nde, Ndonge, which often appear
in tribal names in East Africa. The
Bantu people were not identical to the Zulus
and the Kafrid lakeside fishermen but remained
faithful to their frugivorous subsistence. Their
western branch settled down in West
Africa and may be identified
with Guineids.
The tribal structure of African blacks may be reconstructed according
to frequency in chains of ethnonyms and linguonym. Table 1 takes efforts
to render a rough subcategorisation of African
tribes without pretending an exhaustive depth of taxonomic considerations.
Its right-hand column suggests frequency rates ruling among three principal
branches of black people. The main conclusion is that Oldowans
lacked a permanent tribal ethnonymy because the
African, Melanesian, Australian, Chinese and Latino-American Negrids bear different tribal names. African blacks
created their tribal ethnonyms later in the era of
the Sangoan culture.
|
Negrids →
Oldowan Palaeo-Negrids
+ Sangoan Neo-Negrids
Sangoan Neo-Negrids → Congids
+ Guineids + Bantuids
Congids (Central Africa) → Mbangwe-Ngom
tribes (Y-hg E-M96, mt-hg
L0):
Mbangi 2´, Mbangwe 2´, Ngom 2´, Ndasa 1´.
Guineids (West Africa) → Mbum-Gbaya (Y-hg E1a-M132, mt-hg L1, L2):
Mbum
7´, Mbonga, 1´, Gbaya 8´, Gbagyi 3´.
Bantuids (East Africa) → Amba-Ndonge tribes (Y-hg E1b-P177, mt-hg L0a, L0d):
Ndogo 1´, Ndo 3´, Nding 1´, Ndrule 2´, Amba 3´.
|
Table 1.
The division of African Negrids in the light of ethnonymic
routes
African Dolichocephalic Races
The ethnic dominant in the African
continent were the Bantu Negrids, who
preferred vegetal food and were confined to the ecotype of damp humid
rainforests. These predispositions specialised them as plant-gatherers and
banana-eaters. In the Neolithic such (pre)agricultural inclinations turned
them into slash-and-burn farmers. Now they
are classed as Negrids, Negroids,
Congoids, Congids or Congolids. The terms of blacks, Negroes, Melanodermi or Melanochroi
are applied also to other African races although the criterion of skin
pigmentation is often superficial and misleading. It is a secondary trait due
to subsequent hybridisation because the primary racial phenotype of black
races is determined by their skeletal osteology and
craniology. They belong to the lineage of Jolly’s robust herbivores with vegetal subsistence,
agricultural dispositions and axe-tool industry used for digging out plant
roots and slashing woody species. They were accustomed to upright gait and
nomadic life in clearings of bamboo rainforests in the tropical equatorial zone.
The rainforests of Central Africa served as the
original cradle-land of all Negrids. They stemmed
from the Lower Palaeolithic Oldowan culture of Homo
erectus and preserved their ethnic and cultural core in ideal unimpaired
conditions. Their offshoots encompass all dark-equatorial racial complexes
surviving in the equatorial zone but they include also hybrid varieties of depigmented white-skinned races into the colder
subtropical, boreal and arctic areas of Eurasia. Their
differential analysis is illustrated by Mp 3 and Map 13 entered in the first
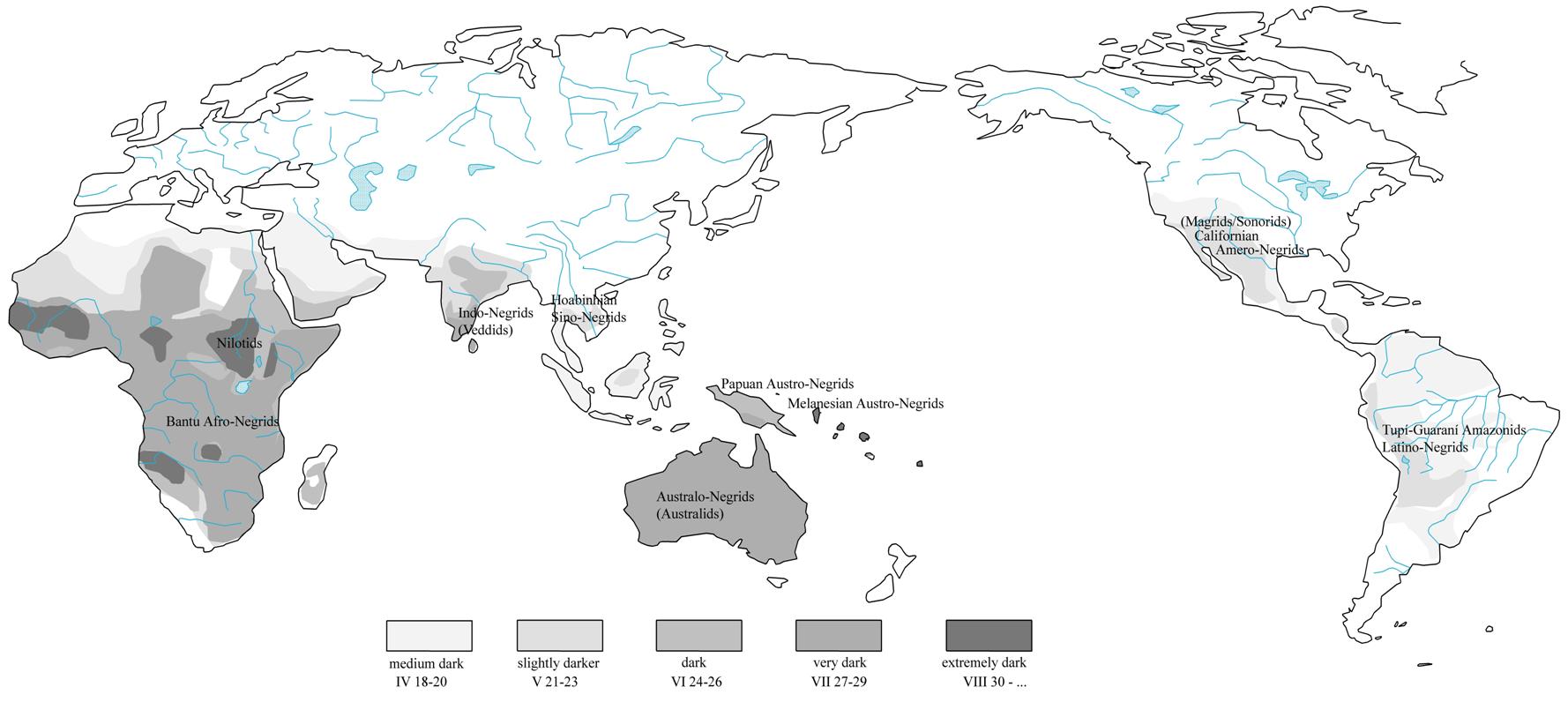
volume. Their purpose is to contrast the height of human stature in Negrids to other anthropological varieties of Africa. Map 3
reproduces the background of Renato Biasutti’s map of African anthropology enriched by colours that outline the
approximate distribution of racial complexes responsible for such indices.
The black Negrids and their equatorial race engendered more than
one third of humankind that was predestined to plant-gathering, farming and
manufacturing hand-axe tools. A half of their populations abandoned tropical
regions and colonised the Eurasian continent with colder climate. Their
cultural morphology shows derived metamorphosed patterns caused by
interbreeding with Altaic hunters. Notwithstanding, their assimilative impact
could not uproot their filial allegiance to genetic axe-tool traditions. This
is how the axe-tool makers have formed a compact
group of cultures with tall stature, prominent dolichocephaly
(long heads), hypsicrania (tall skulls), euryprosopia (large broad faces), platyrrhinia
or chamaerrhinia (broad noses), brachycormia
or metriocormia (shorter or medium-size trunks) and
macroskelia (long legs, long lower extremities).
Their
Eurasian, Siberian, Indic and North American splinters lost much of this
genetic dowry. They were all children of the new cultural Acheulean
cultural unity that struck roots in South Arabia. Their
stock stemmed from Caucasoids, whose pigmentation
made Huxley classify them as brown-skinned melanochroi.
They grew into the main Asiatic branch of Caucasoid axe-tool cultures and
later also Elamitoid agricuturalists.
Independent development afflicted their westward-oriented branch that gave
birth to Anatolids, Danubian
Europoids and Scandinavian Nordids.
These racial offshoots underwent a large-scale depigmentation
of skin, hair and eyes. However, interbreeding with Levalloisian and
Mousterian flake-tool cultures made them adopt also their leptoprosopia
(narrow faces) and leptorrhinia (narrow noses).
|
The Anthropogenesis
of Negrids
All ethnic families are
interrelated with our remote forefathers, who descended from the equatorial
race of African Negrids. Their stock encompasses
almost half of humans stemming from prehistoric axe-tool makers, plant-gatherers
and preagriculturalists. Table 2 depicts their
evolutionary splitting by means of a genealogic tree graph pursuing the
branching of Y-DNA haplogroups (their abbreviations
are written E-hg, I-hg etc. This graph omits the lineages of Lappids, Scytho-Ugrids and
Ural-Altaic flake-tool makers and concentrates only on the evolution of
equatorial dark-skinned Negrids, who mixed with
northern boreal races and gradually developed into light-skinned Caucasoids and Europids.
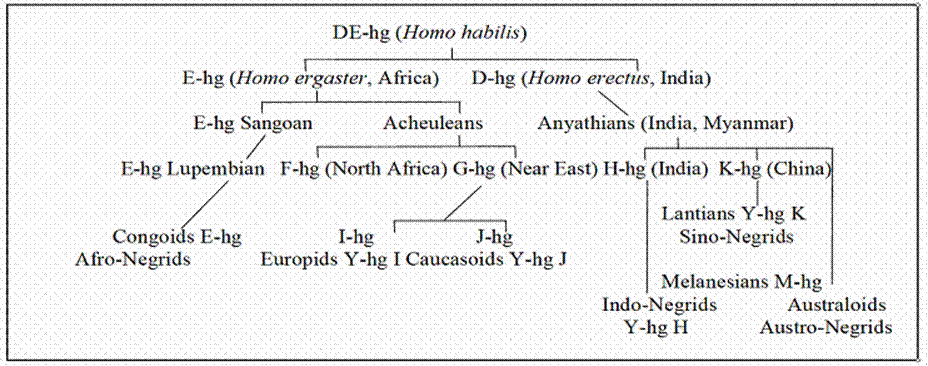
Table 2.
The phylogenetic tree of
plant-gatherers and axe-tool makers
The forthcoming Table 3
attempts to record the parallel splitting of human stocks by the notation of generative grammars. It copes with
several unsolved incongruous discrepancies concerning the haplotypes
K, D and M. It revives several seemingly obsolete terms of archaeology such
as Kafuans, Chelleans, Abbevillians, Anyathians and Campignians classified as Littorids.
They are regarded as outdated but appear necessary for filling up certain
empty pigeon-holes in the evolutionary process. The category of Acheulean culture covers a period that is too large to
express subtle nuances of cultural growth. Archaeologists should follow Louis Leakey,
who specified eleven evolutionary stages of
the Chelleo-Acheulean ‘hand axe culture’.
The chief problem has to do with the descendants of the Y-DNA haplogroups DE and D. They headed for India, China and Melanesia and must have
participated in the birth of Australoid races. A
large gap divides from the Oceanic haplotypes M and
S, whose rise is erroneously associated with the genome K of Europoid Littorids spreading
the shell-midden Lapita
culture (5,000 BC). In spite of a few unclarities,
Table 3 gives an approximate but instructive visual representation of progress
in the largest stock of humanity.
|
Archaeological axe-tool cultures
|
Dolichocephalic
anthropological groups
|
Y-DNA
population genetics
|
|
Kafuans ® Kafuans + Oldowans
|
H. ergaster ® H. ergaster + H.
erectus
|
DE ® DE + D
|
|
Kafuans ® Kafuans + Anyathians
|
H. ergaster ® H. ergaster + Indo-Negroids
|
DE ® DE + F
|
|
Kafuans ® Kafuans + Sangoans
|
H. ergaster® Paleo-Negrids + Neo-Negrids
|
DE ® DE + E
|
|
Kafuans ® Sangoans + Chelleans
|
Paleo-Negrids ®
Neo-Negrids + Chelleans
|
DE ® E + F
|
|
Sangoans ® Epi-Sangoans + Lupembans
|
Neo-Negrids ® Congoids + Bantuids
|
E ® E1 + E2
|
|
Chelleans ® Chelleans + Abbevillians
|
Chelleans ® Chelleans + Caucasoids
|
F ® F
+ GHIJK
|
|
Abbevillians ® Acheuleans + Irrawaddians
|
Caucasoids ® Gothonids + Burmids/Hmongids
|
GHIJK ®
HIJK + G
|
|
Acheuleans ® Micoquians + Yabrudians
|
Gothonids ® Europids + Hethoids/Elamitoids
|
HIJK ® IJK + H
|
|
Micoquians ® Macrolithic + Campignians
|
Europids® Gothids+Jomon/Vindhya Littorids
|
IJK® IJ
+ K
|
|
Macrolithic ® Gothids + Gothonids
|
Gothids ® Gothids + Levantine Hethoids
|
IJ ® I + J
|
|
Gothids ® Corded Ware + LBK Danubians
|
Gothids ® Gotho-Frisians + Langobards
|
I ® I1
+ I2
|
|
Epi-Oldowans ® Hoabinhians + Lapita?
|
Epi-Oldowans ® Papuasids + Melanesids
|
K2bl ® S
+ M
|
Table
3. The branching of dolichocephalic
cultures, races and haplogroups
Extract from Pavel Bělíček The Differential Analysis of the Wordwide
Human Varieties. Prague 2018, pp. 11-14
|
|