|
|
|
|||||||||||||||
|
|
|
|||||||||||||||
|
|
|
|
||||||||||||||
|
|
|
|
||||||||||||||
|
|
|
|
||||||||||||||
|
|
|
|||||||||||||||
|
|
|
|
|
|||||||||||||
|
|
The Racial and Linguistic Groups of Negrids,
Bantuids, Melanids and Amazonides Clickable terms are red on the yellow background |
|
||||||||||||||

|
Table 1. The Systematic Glottogenesis of Human Language
Families |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Map 1. The Migration Routes of Bantu Negrids to Papua, Melanesia and Amazonia |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The Oldowan Colonisation
Traditional anthropology operates with
theoretical models of the triumphal spread of Homo sapiens and his
out-of-Africa propagation.1 It is
based on a pliable all-explaining construction that unfortunately lacks
unequivocal support in archaeological evidence. Its argumentation buries Homo
erectus as an extinct species and replaces his reign by a gracile successor inhabiting lakeside districts of the
East African Depression. These territories were formerly occupied by his
rival Homo rudolfensis indulging in
waterside ecosystems. His life-style was probably inherited by the race of
East African Kafrids producing Levalloisian
flake-tools. About 500,000 and 125,000 BP they set out on several migrations
heading for There exists a grave suspicion that their
gentle traits misled anthropologists into considering them as the sole
forefathers of all human races. They did not realise that Classic comparative linguistics
presupposes more conservative dating, the founder of Bantu philological
studies Harry H. Johnston advanced much later estimates of Bantu origins. He
presupposed that obvious analogies between African Bantu tongues and
Australian Aboriginal had been due to contacts dated to a few centuries AD.2 Anthropologists attempted to solve
this crux by searching for the earliest African digs and fossils. For a long
time they recognised the earliest ancestor of black Bantu people in the
African fossils of Asselar man. His remains were
found near The issue of common ancestry linking
African, Melanesian and Australian black people into one anthropological
group is still open to dispute. The prevailing opinion regards their racial
groups as independent outgrowths of Homo sapiens that developed on
different continents by adapting to similar tropical climate. Our opinion
proclaims that they represent the progeny of the earliest Oldowan
colonists. The
chief objections to the concept of Palaeo-Negrid
unity emphasise great differences in hair. The hair of Melanesians is wavy or
straight owing to contacts with neighbouring Polynesians. The hair of Australids is as wavy, silky and kinky as
that of Veddoids. The original racial archetype is preserved only in
African Negrids, whose hair is woolly, frizzly and curly. Racial differences among black melanodermic and ulotrichous
types are mostly due to secondary influences but they also disclose common
archaic traits receding in African archetypes. For instance, Australids differ from African Negrids
by higher occurrence of the so-called ‘sagittal
keel’. Originally, it was typical of Homo erectus as a survival of crista sagittalis
in Paranthropus boisei.
Other archaic features of Australids include also
greater prognathism and large heavy jaws betraying
vegetal subsistence. Such findings do not contradict common origin but
confirm the original unity by preserving the more archaic status. They prove
that the original lineage of robust herbivores may have survived better in
remote isolates. The progeny of Homo erectus and Oldowan wanderers appeared also in Amazonian peasants
pertaining to the Tupí-Guraní language family.
Their burial customs prescribe excarnations of the
dead grandfathers’ skulls and skeletons. Formerly, their tribes practiced
ritual cannibalistic endophagy, their
survivors originally ate their flesh and drank their blood in hope they would
take over their magic spiritual powers. Their recent progeny replaced these
barbarous ceremonies by consuming them in burnt ashes baked in cakes or
blended in drinks. The Bantu tribe Geshu boil their dead by night, cut off pieces of the
flesh, divide them among women and leave the rest to the jackals.1 This happens also in the The Amazonian connection of Palaeo-Negrids is scented also by geneticists.4 An important
observation was made by Christy Turners,5 who studied dolichocephalous crania of Amazonian
tribes in The undoubtable
existence of prehistoric Palaeo-Negritic unity
pulls down all the walls of traditional prehistoric studies. The American
linguist Johanna Nichols applied statistical methods to prove that vocal languages
started diversifying in human species at least 100,000 years ago.8 Comparisons of Bantu, Australian,
Melanesian and Amazonian languages bring evidence of many structural
analogies. They show that it is difficult to find common lexical roots in
their word stock, but there exist conspicuous parallels in categories of
phonology and grammar. These languages include initial prenasalised
stops
There exist grave reasons for acknowledging four principal palaeo-races that stood at the cradle of humankind and
arose by adapting to the basic ecotypes in (i) the herbivorous
plant-gatherer Homo erectus remarkable for the Oldowan
culture (Y-hg E), who may have become the progenitor of the equatorial race
of Negrids, (ii) the carnivorous
hunter Laetoli man noted for volcano burials
and the beehive-dwelling culture of Eickstedt’s Khoids with the haplotype
Y-hg B, (iii) the piscivorous fisher Homo rudolfensis
with the Y-hg B, who developed the Levalloisian culture (500,000 BP) and gave
birth to the race of lake-dwelling Kafrids in the
East African Depression, (iv) the omnivorous
and insectivorous race of Eickstedt’s Sanids affiliated with the Negrillo
and Lappids. These
lineages ascertain the genetic continuity of certain ecotypes and modes of
subsistence between hominids, hominins and human
races. They may have been responsible for the origin of several survivors
differentiated into robust plant-eaters (Negrids),
dwarfish omnivores and insectivores (Sanids), lacustrine fish-eaters (Tungusoid
Kafrids and the carnivorous hunters (Coon’s Capoid race, Biasutti’s
Steatopygids9, Eickstedt’s
Khoids). The Analytic Decomposition of Cordal
Axe-Tool Languages
It is
erroneous to identify the birth of Indo-European with Old Indian and imagine
its glottogenesis as a series of sound shifts
transforming Vedic Sanskrit phonemes. Jacob Grimm’s laws overestimated
Sanskrit as direct evidence of the early state of Indo-European and traced
its development as a westward move from
Table 10. Conservative embeddings and
regressive intrusions in Old Indian consonantism The first autochthons in The first Indo-European
newcomers were Campignian Littoralists
(10,000 BC) with cordmarked pottery, who colonized
the The Dravidian element in Old
Indian was represented by retroflex consonants written as ṭ, ḍ, ṇ,
ṣ, ẓ, ḷ, ɾ̣, ɹ̣
but the IPA standard records them as /ʈ, ɖ ,
ɳ, ʂ, ʐ, ɭ, ɻ, ɽ/. They were notable
for pronunciation with the tip of the tongue bent backwards in a concave or curled shape. Their use was obliterated in
most language families but their remains often survive in the affricates tr- dr-, tl- and dl-. Most types of notation
do not distinguish their apical and laminal
pronunciation. The laminal retroflex consonants
were characteristic of Tungusoid fishermen, who
disseminated them in One of typical Indo-European consonantal
clusters is seen in clusters combining sibilants with plosives and sonants. Their majority was formed by the concatenations sp-, st-, sk with presibilised surd plosives and the groups sn-, sl-, sr- formed by presibilised surd sonants. The fact that they were derived
from surd sonants betrays their non-Indo-European
origin because the native IE sonants were voiced. The residues of presibilised clusters appeared also in Georgian and
Caucasian languages, which suggested a false impression of their close
genetic affinity. In fact, they arose as an Europeanisation
of Altaic roots with fortis and glottalic
stops. Their original fortis pronunciation was
perceived as monophonematic aspirated stops
ph- and hp- or a pair of two
phonemes ph- or hp-. A similar process of deaspiration took place
in Czech chvíle ‘while’ derived from Old High German hwila and New German Weile.
Such assibilation was common especially in assimilating the glottalic ejectives of Scythoid kurgan-builders and Khoisan
herders with copular beehive dwellings and leaf-shaped lance-heads. The South
African Khoekhoe tribes belonged to the group of pastoralists such as the Maasai,
Musgu, Berber Imazhigen
and Iberian Basques, whose ethnonyms resonate also
in the place names Altaic intrusions into Indo-European encompassed also retroflex plosives that are usually transcribed in India as /ṭ, ḍ, ṇ, ṣ, ẓ, ḷ, ɾ̣, ɹ/̣ but in the IPA notation they are rendered as /ʈ, ɖ , ɳ, ʂ, ʐ, ɭ, ɻ, ɽ/. These sounds come from Turcoid and Tungusoid languages and often underwent dephonologisation into r-affricates or l-affricates. Such assimilative decomposition led to r-affricative biphonemes /pr, tr, dr, kr, mr, nr/, or l-affricates /pl, tl, dl, kl, ml, nl/. The final stage of dephonologisation turned fortis voiceless consonants sounds into presibilated consonantal clusters. Fortis retroflexed cacuminals were decomposed into cacuminal clusters /spr- str- skr-/, while fortis retroflexed laminals degenerated into laminal clusters /spl- stl- skl/. Almost all reconstructions
of Indo-European count with a special series of palatals /*k', *g', *g'ʰ/ transcribed also as /*ky, *gy*,
gyʰ/. Indian palatal phonemes have
to be classified as an import of the short-sized Negritos,
Alpinids and Lappids, who
detested velar and guttural consonants and removed them by satemisation. The first wave of Negritos
arrived in The Scythoid
element was present in |
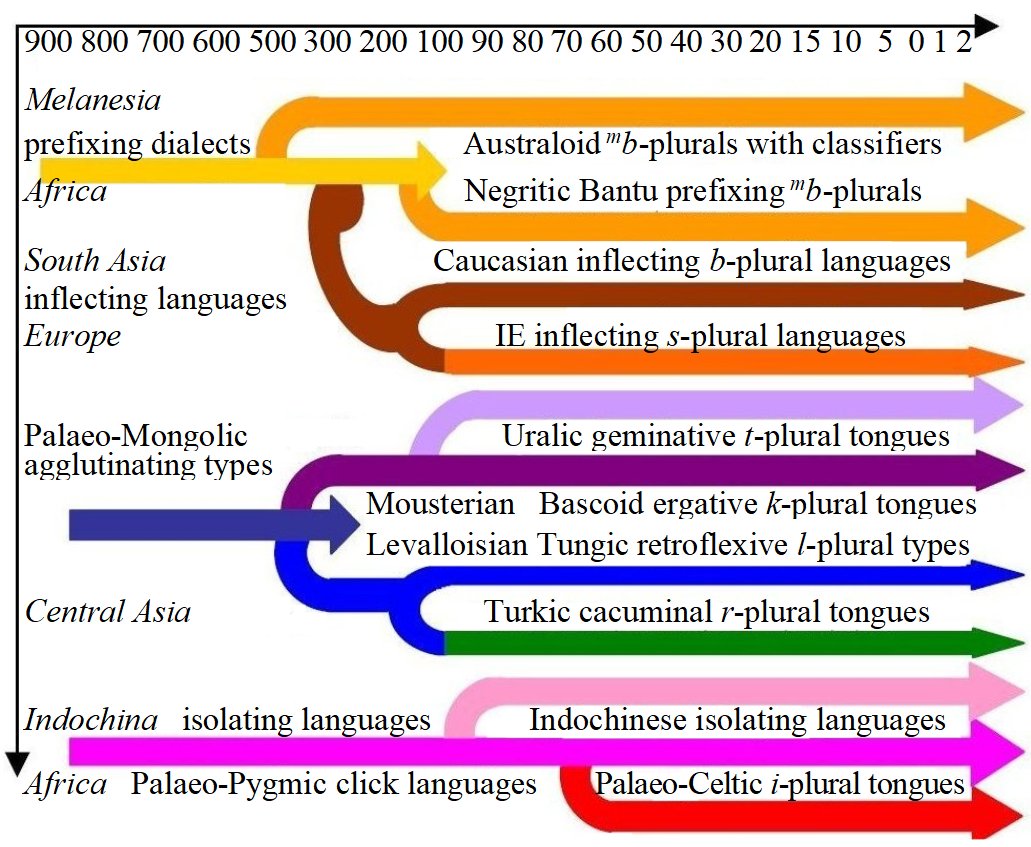
The Palaeo-Negric, Palaeo-Hmongic, Palaeo-Melanese and Palaeo-Amazonian
Phonology There
were several principal turnabouts in the tongues of robust dolichocephalous
axe-tool makers. They were all herbivorous plant-gatherers and seed-eaters,
who dug up their crops with pebble-stone-choppers and hand-axes. Their
eastern migrations headed for the The
original speech patterns of the black Negrids and Melanids were preserved best in Bantu languages. When the ancient Oldowans
moved to In The problem of Acheulean,
Caucasian and Indo-European continuity is solved by focusing on avalanches of
ephemeral sound shifts instead of considering genetic stability and
structural typology. The white Europoid race is an
outgrowth of macrolithic hand-axe populations of
tall dolichocephals with vegetal subsistence and
(pre)agricultural dispositions, and its development competed with the Altaic
races with flake-tool cultures. These cultural traditions did not differentiate
by unilinear monogenesis from a common prehistoric
unity but pursued independent growth by paragenesis
in several interfertile and interbreedable
racial lineages. Their archetypal differences were manifested by absolutely
incompatible phonologies and grammatical systems. African Negrids,
Asiatic Caucasoids and European Nordids
applied cordal languages, whose consonantism was based on the vibration of vocal cords
and the opposition of voiced and surd phonemes (Table 5). They pronounced
vocalic cordal phonemes based on open syllables and
phonemes produced by airstream passing through
vibrating vocal cords. If there were any structural changes, they were caused
by mixing with Altaic agglutinating systems.
Table 5. The cordal
phonology of dolichocephals with macrolithic hand-axe industry The Situation of Indo-European
among Grammatical Systems
Classical historical grammar fulminated
thousands of sound shifts but brought few testimonies of similar radical
changes in grammar, accidence and syntax. Phonological repertories tend to
exhibit radical innovations but they remain unparalleled in other linguistic
aspects. In cultural evolution most grammatical categories remain relatively
untouched and adopt mutations only in suffixal
morphemes. This discrepancy is removed if linguistic analysis focuses on
spoken dialects instead of the official written standard of national
languages. This is another good reason for promoting linguistic typological
studies that do not restrict their focus to describing individual language
structures but build bridges between tongues with similar types of traits.
Table 8.
The nominal morphology of dolichocephals
with macrolithic hand-axe industry Elementary grammatical systems fall into
three types of nominal and verbal morphology. The gender-oriented morphology
is attributable to the language family of tall dolichocephals
with hand-axe industry and vegetal subsistence. In its original appearance documented
in African, Melanesian and Australian Negrids it
partitioned nouns into classes of animate, inanimate, vegetal and arboreal
classes. These classed were distinguished by prefixes put in front of nouns.
In the Horn of Africa their family ran upon Asiatic races with agglutinating
language structures and transitioned to suffixing morphology of inflecting
type. The group of Asiatic plant-gatherers, hoe-cultivators and
agriculturalists reduced the system of twelve nominal classifiers to the
opposition of animate and inanimate nouns. Their category included humans,
animals, animistic spirits as well as sacral deities. This
categorisation survived also in Anatolian tongues until their further
expansion in the Balkans encountered Gravettian
tribes of Alpinids with sex-based gender
classifications. Their clash resulted in the rise of sex-based nominal gender
enriched by masculine o-stems and feminine a-stems. The core of
European Gothids accepted the dual opposition of
masculine and feminine gender but their core remained reluctant to their
addition and continued to adhere to nominal i-stems.
Their subclasses coexisted with Caucasoid vegetal u/w-stems
that can be explained as remains of Caucasoid b-plurals referring to
agricultural crops and instruments of farming activities. The classification
of Indo-European thematic and athematic stems may
be regarded as a hold-over of ancient invasions and infiltrations surviving
in residual form in the territory of (from
P. Bělíček: The Analytic Survey of European Anthropology, Prague 2018, p.
35-42) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||