|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
The Y-DNA Haplogroups of Principal Races Clickable terms are red on the yellow background |
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Table 2. The
Hypothetical Model of Long-Range Genetic Transitions of Y-DNA Haplogroups |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Map 1. The World Ethnic Bearers of Y-DNA Haplogroups
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Table 3. The branching of dolichocephalic cultures, races and
haplogroups |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
The Mid-Range Reach of
Chromosomal Genome Typology
Current population genetics defends African
origins and Chris Stringer’s
‘single-origin model’. It supports theories of strict monogenism in belief
that one sapient race colonised the world from one homeland in
Table
4. The simplified genealogic tree of Y-DNA haplogroups The out-out-Africa wandering
sounds like as tenable hypothesis but its dating must be shifted to the
earlier times of Oldowan migrations from 1.8 mya to 0.5 mya. There exists
little evidence supporting the alternative theory of an out-of-Asia model but it is worth taking
into account. Besides the Riwat flake-tool culture (2 mya) in north Pakistan
it is necessary to scrutinise Asiatic hominins as an independent genetic
strain evolving inclinations to green, yellow and red pigmentation. Asiatic
orangutans and gibbons exhibited green, grey, yellow and red hair
pigmentation. Prehistoric megalith-builders in According to temporal
durability, anthropological parameters may be divided into short-range
markers, mid-range markers and long-range markers. The ABO blood groups
belong to the category of long-range markers because they encroach upon
typology shared by leptorrhinian apes in the Most anthropologists adhere
to the Chris Stinger’s monogenetic single-origin theory that derived the
descent of humankind from African hominids. They support the out-of-Africa
model of anthropogenesis that claims that all living races owe their roots to
Homo sapiens born in eastern Current accounts of population genetics
have summarised the monogenetic classification that endeavours to arrange all
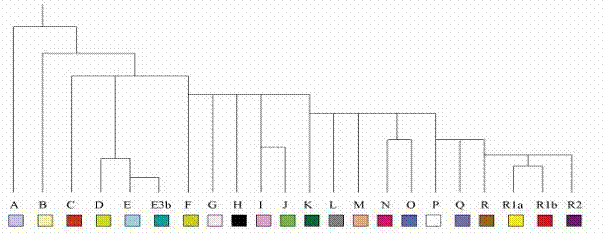
human chromosomal Y-DNA haplotypes into one evolutionary tree. It attempts to
explain their rise via series of successive mutations. The contemporary
widely-accepted ordering of Y-DNA genomes is outlined in Table 4. It enjoys
an autoritative influence but cannot conceal some apparent inconsistencies.
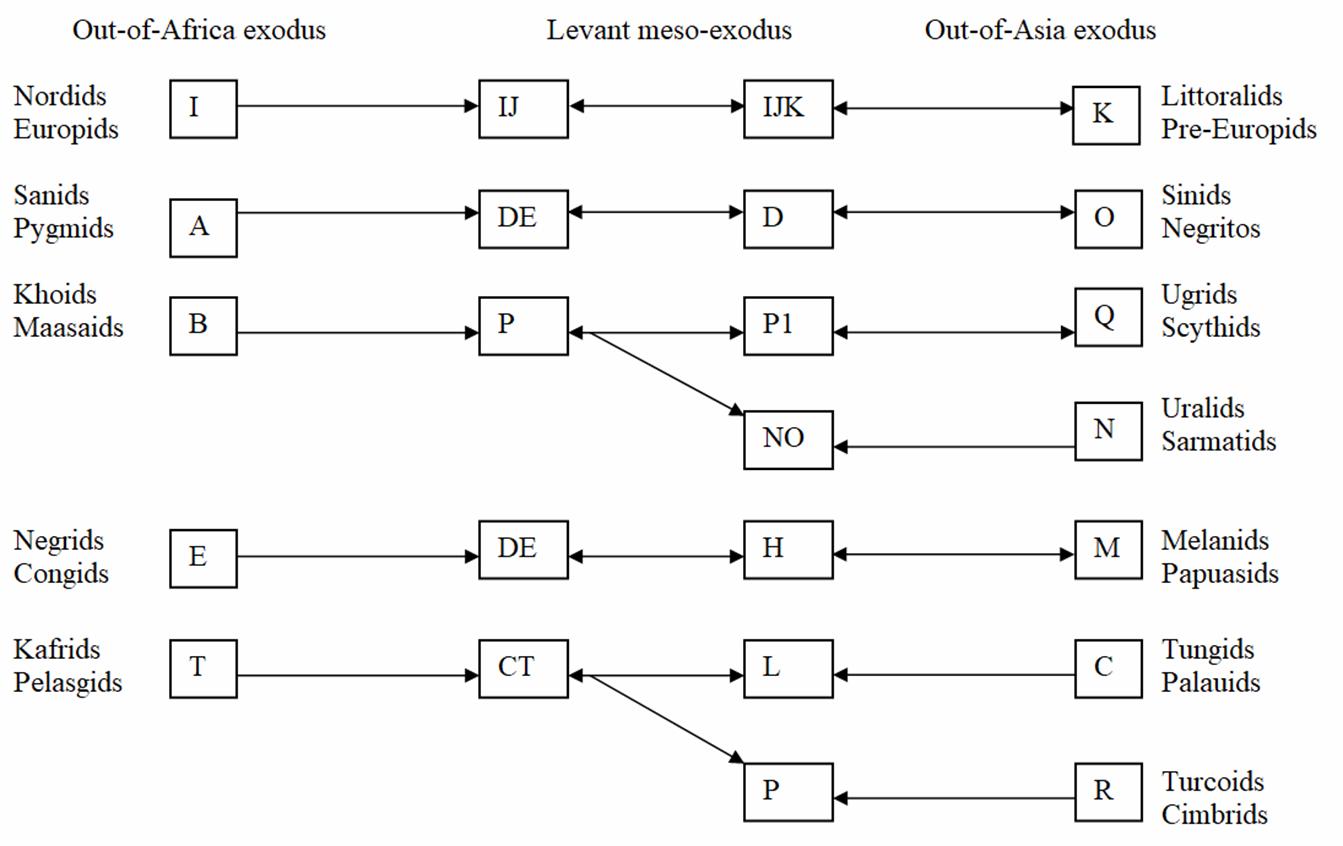
It fails to associate the African Negrids with the Oceanic Melanids and the
African Pygmids with the Negrito of South East Asia. Neither does it
associate the Uralids with the haplotype N with the Ugrids who share the
Y-haplotype Q with Amerindian wigwam-dwellers. To our surprise, it
finds the N-haplogroup of Uralids closely related kinsmen in the shortsized
Sinids with incompatible isolating languages. Table 3 takes these
incongruences into account and proposes a hypothetical model of genetic
paragenesis between interbreedable racial phenotypes with a high degree of
mutual interfertility. Extract from Pavel Bělíček: The Synthetic Classification of Human
Phenotypes and Varieties. Prague 2018,
pp. 19-20 |
|
The Anthropogenesis of the Dolichocephalous Axe-Tool
People
All ethnic families are
interrelated with our remote forefathers, who descended from the equatorial
race of African Negrids. Their stock encompasses almost half of humans
stemming from prehistoric axe-tool makers, plant-gatherers and
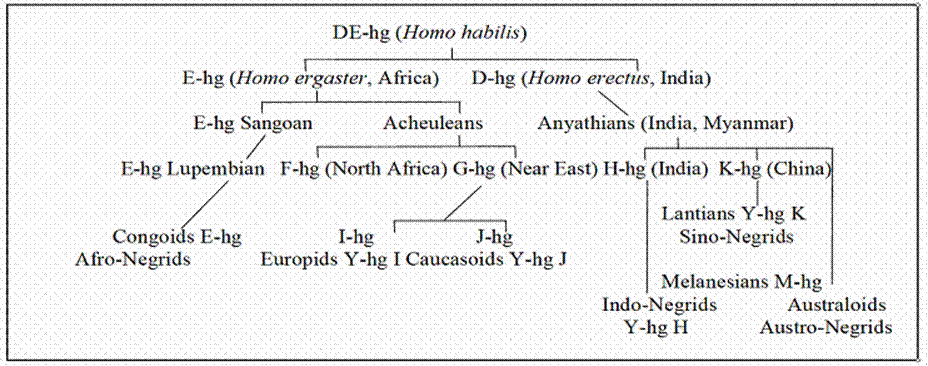
preagriculturalists. Table 3 depicts their evolutionary splitting by means of
a genealogic tree graph pursuing the branching of Y-DNA haplogroups (their
abbreviations are written E-hg, I-hg etc. This graph omits the lineages of
Lappids, Scytho-Ugrids and Ural-Altaic flake-tool makers and concentrates
only on the evolution of equatorial dark-skinned Negrids, who mixed with
northern boreal races and gradually developed into light-skinned Caucasoids
and Europids.
Table 5. The phylogenetic tree of plant-gatherers
and axe-tool makers The forthcoming Table 3 attempts to
record the parallel splitting of human stocks by the
notation of generative grammars. It copes with several unsolved
incongruous discrepancies concerning the haplotypes K, D and M. It revives
several seemingly obsolete terms of archaeology such as Kafuans, Chelleans,
Abbevillians, Anyathians and Campignians classified as Littorids. They are
regarded as outdated but appear necessary for filling up certain empty
pigeon-holes in the evolutionary process. The category of Acheulean culture
covers a period that is too large to express subtle nuances of cultural
growth. Archaeologists should follow Louis Leakey, who specified eleven evolutionary stages of the Chelleo-Acheulean
‘hand axe culture’.1 The chief problem has to do with the descendants
of the Y-DNA haplogroups DE and D. They headed for Extract from Pavel Bělíček:
The Differential Analysis of the Wordwide Human Varieties. Prague 2018, pp. 11-13 |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||