|
|
The prehistoric Oldowans did not have their stable clan organisation and
more permanent tribal ethnonymy. As a consequence, they did not leave durable
vestiges of ethnic clan hierarchy in their place names and it is not possible
to find common roots of their phratries. The Congids display a lot of
toponyms with the root Cong- and the Bantuids abound in the tribal
name Nde. The ethnic label of the Bantu probably arose by adding the
animate plural prefix ba- to the root Nte or Nde.
Chinese Sino-Negrids later produced their own independent ethnonymy several
thousand years ago. The Neolithic changes welcomed them as Hoabinhians but
now they survive as the Hmong-Mien stock. American Negrids were remote
descendants of Chinese Homo erectus lantianensis but did not inherit
his ethnonymy. Now they chiefly distinguish the Tupí and Guaraní branch of
their family and these ethnonyms cannot be older than 10,000 years.
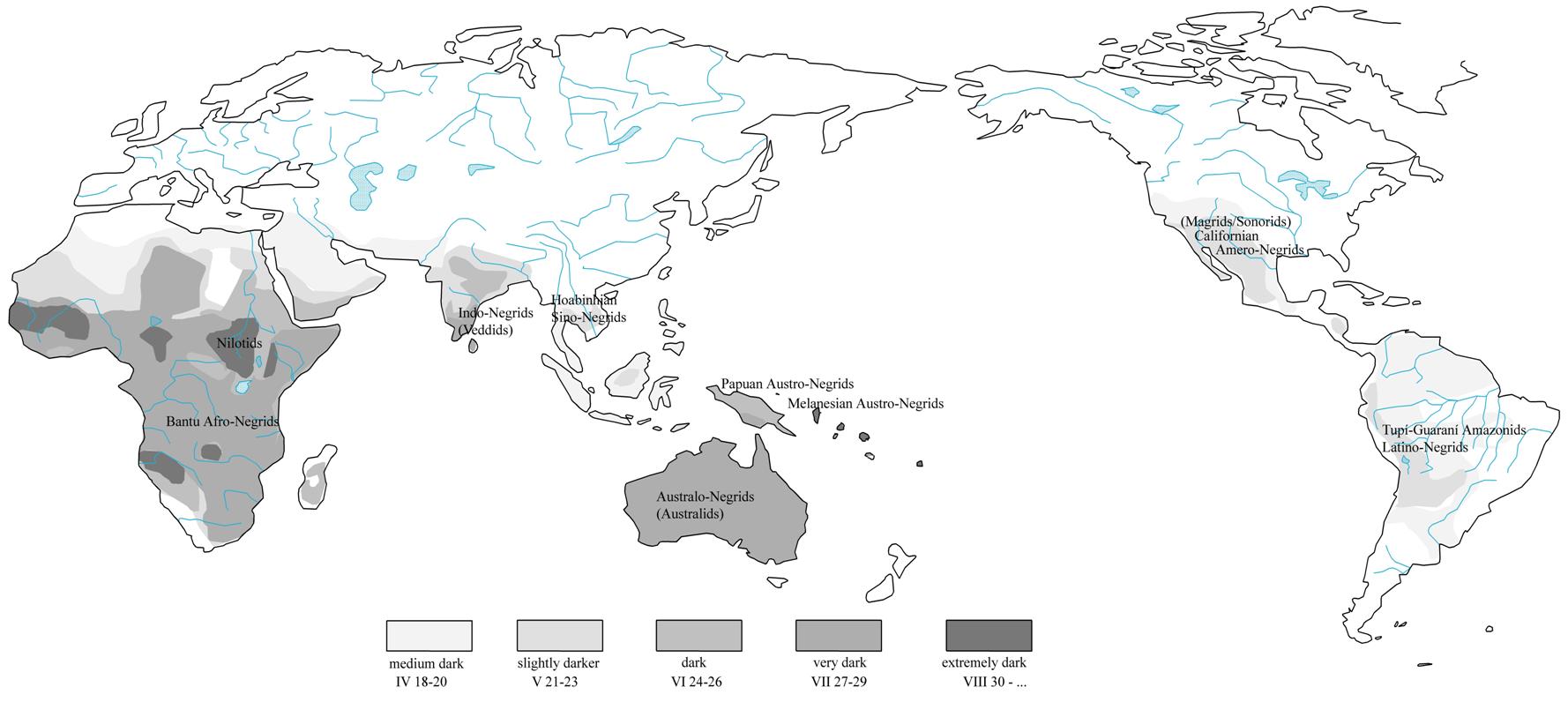
Melano-Negrids (cca 30 000
BP): extreme archaic platyrrhinia, prognathism, dolichocephaly, residual crista
sagittalis characteristic of Homo erectus; ethnonyms with prenasalised
stops mb-, nd-,
ng-, which stand in correlation to voiced consonants.
Australo-Lappids, Negrito Sinids,
Lake Mungo site (Negrito diaspora, 56,000 BC):
short-sized stature with short lower extremities, brachycephaly, pearl shells
as pubic covering called jakuli or riji in Baada, bead-belts
with pubic shell and meander-like key-pattern ornamentation; humpy lean-to,
windbreak shelters, cremation burials; reduplicative morphology, Windowinda, Wembawemba, Wandandian, ethnonymic roots Kal-/Gal-, Wend-/And-/Ind-.
Oceano-Tungids (30 000
BP): a continuation of Austro-Tungids such as Telugu, Tulu in Dravidian
India, tribes speaking the Formosan languages of Taiwan and Tagalog on the
Philippines related also to Malaysian, Indonesian and Hawaiian; ethnonymic roots Bal-,
Tung/Dan-, Ker-; atlatl spear-throwers.
pit-graves with ochre burials, lakeside ecotype, archaic proto-tepee huts,
conical round lodges from crossed poles; gracile slender body-build, Y-haplogroups C4-M343, 60.2 %, C*-M130, 2.0 %.
Oceano-Turanids (cca 30 000
BP): Y-haplogroups R1-M173, circumcision,
knife-throwers, sickle-shape boomerang throwing, loincloth, cave-paintings,
cave shelters with Azilian-like imprints of phalanges, phallomorphous shamans
with herds of wild small game, X-ray representation of killed animals with
translucent intestines for hepatomancy and divination from livers.
Oceano-Scythoids (cca 30 000
BP): a continuation of Munda beehive-dwellers with mummification and burial
rock-pile mounds in India, Aranda people and ethnonymic roots in Manda-,
Yurak-.
Oceano-Littoralids (cca 30 000
BP): shell midden cultures, littoral beachcombers, Lapita culture,
Y-haplogroup K*.
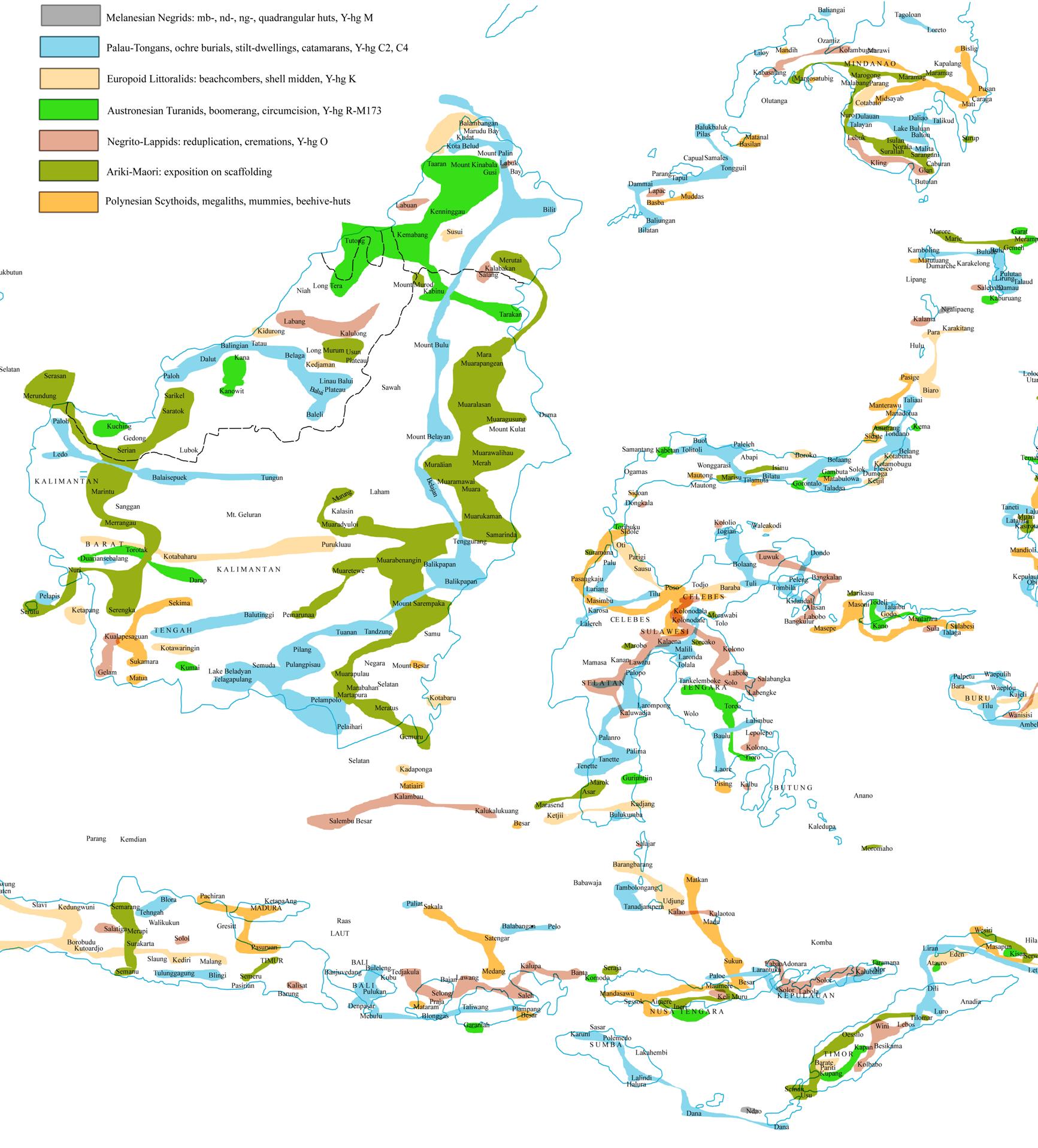
Table 1. The analytic
typology and taxonomy of Aboriginal Australians
The ethnonymy of Australian
black men does not depend upon on Hoabinhian sources, either. However, it may
be recognised easily by the bisyllabic relief of clan names with voiced stops
prenasalised the first initial position as well as in the second syllable.
Other ethnic and tribal minorities arrived in Australia
with an accomplished system of tribal organisation and transplanted archaic
Eurasian ethnonyms to the then Sahul and the now Australian continent.
|
The anthropology of
Australian racial varieties cannot rely only on ethnonymy and anthropometric
measurements. It must involve into its scope also ethnographic cultural
ethnology that makes it possible to links Australian Aborigines with their
Eurasian progenitors. This is why Table 10 strives to coordinate racial
indices with ethnic peculiarities in architecture, clothing, divination and
burial customs. The evidenced diversity of the haplotype genetic pool can be
interpreted as a proof of multiple invasions of heterogeneous tribes of
different ethnic origin. When compared to New Guinean explorations,
Aboriginal populations in Australia
bear relatively low percentages of the Melanesian haplogroup M, which must be
due to the dominant superposition of the Australo-Tungids with the C type.
Notwithstanding, judging according to tribal ethnonyms with the prenasalised
stops mb-, nd-,
ng-, they represented a very populous substratum.
In the 1910s the school of diffusionist anthropology
managed to revolutionise the 19th century dogmatic thought by
opposing the popular belief that all continents had one unique ancestor. It
admitted migratory diffusion and refuted insisting on theories that all
continents had been populated by one member of Homo sapiens, who
fathered all local subspecies. The comparative evidence of archaeology
demonstrates that all continental races had obvious typological parallels in Eurasia
or Africa. It is much more probable that Oceania,
Polynesia, Australia
and America
were visited by several diverse types of archaeological cultures, whose
bearers exhibited incompatible racial traits. Despite subsequent secondary
acculturation their archaeological complexes bore evident resemblance to the
well-known human stocks of Eurasian and African prehistory.
|
Ecotypes
|
Anthropological and cultural traits
|
|
herbivores
ulotrichous
chamaerrhines
|
genuine Australids or Australo-Negrids with the ABO group O and
Y-DNA haplogroup M, plant-gathering and vegetal subsistence, dolichocephalous
skull with broad platyrrhine noses and high nasal indices, prenasalised
stops
|
|
Lacustrines
lake-dwellers
ichthyophages
|
Australo-Tungids, riveside and lakeside ecotypes, nomadic fishermen,
round conical tents with crossed poles, pit graves, ochre burials, the
dead corpses are besprinkled with
red hematite paint, Y-DNA haplogroups C4 and C2
|
|
Troglodytes cave-dwellers
ichthyophages
|
petroglyphs, rock painting with imprints of phalanges, roentgen
drawings of animals for hepatomancy and haruspicy from the entrails killed
game, curved boomerangs and knives used as weapons, archaic
Y-haplogroup R1-173
|
|
Littoralists
beach-combers
|
shell midden, sand-dune dump of kitchen waste, rectangular
monopitched huts
tall stature, dolichocephalous skulls, low head indices
|
|
Mummifiers
beehive dwellers
|
beehive huts in circular campsites, the dead corpses are anointed and wrapped in long
pieces of cloth, graves are covered by piles of stones, Y-haplogroup Q
|
|
Insectivores
incinerators
|
lean-to summer huts, semidugout dwellings, short stature, Negrito
constitution, brachycephalous heads, curly ulotrichous hair, cremation
burials
|
Table 2. The systematic classification of
Australian aboriginal tribes
Human races do not arise in a few hundreds
of years, their germination lasts more than tens and hundreds of millennia.
What differs the Australian Aborigines from African Negrids is not a new
racial outgrowth but different rates of the archaic Oldowan heritage
preserved in their blood. It survived although many traits were partly lost
or receded due to contacts with alien neighbours. The clear and elucidating
implication of the diffusionist philosophy of ethnic groups is that they
inherit one principal paternal strain but blend it with maternal genetic
strains. On one hand, they inherit the genetic pool of the ancestors in their
original homeland, and on the other hand, they hybridise it with traits
acquired in their new outland plantations. As a result, it makes no sense to
invent new names for continental varieties and derive their origin from
subclades of the first colonists. Population genetics confirms that almost
all continents have at least small admixtures of all human haplogroups. No
continent was populated only by one sole race and by one DNA haplogroup.
Continental paradises tend to have one dominant race and unequal rates of
several subdominant assimilated races. The only reasonable anthropological
nomenclature may be devised by compounds indicating reference to archetypal
stocks but comprising also reference to their secondary continental location.
So Australia
is generally recognised as a home of Australids denotable as Australo-Negrids
but its haplogenetic dominant is the Y-DNA type C4 related to C2. This
genetic group is typical of Tungids, the Tagalog in the Philippines
and Polynesian Tongan tribes. So one of the most populous factions among
Australian Aborigines are Australo-Tungid remarkable for pit-grave burials
besprinkled by ochre paint. A convenient proposal of systematic racial
taxonomy of Australian and Oceanic variants is put forward in Tables 9 and10.
Extract from Pavel
Bělíček: The Differential
Analysis of the Wordwide Human Varieties, Prague 2018,
pp. 38-49.
|
|